튜링의 반응-확산 방정식과 몸의 디자인
Phillip Ball, The Self-Made Tapestry, Oxfrd Univ. Press, Chap.4
기린의 목은 왜 긴가? 다윈의 "자연선택"은 이러한 생명체의 형태들에 대해 그럴듯한 답들을 제공해준다. 변이는 무작위적으로 출현한다. 그 가운데 어떤 것은 자손을 남김으로서 다음 세대에 전달되고 또 어떤 것은 자손을 남기지 못해 소멸한다. 다윈에 의하면 기린의 목이 길어진 것은 목이 긴 기린만이 살아남아 그 유전자를 후손에게 전달했기 때문이다. 목이 짧은 기린을 멸종시키고 긴 기린을 살아남게 한 것은 긴 기린에 유리하게 된 자연환경이다. 기후가 건조해져 초목이 말라가고 관목의 높은 가지의 잎만이 먹을 수 있는 유일한 먹이라고 생각해보자. 목이 짧은 기린은 먹이를 얻지 못해 굶어죽지만 긴 기린은 관목의 잎을 뜯을 수 있기 때문에 살아남을 수 있다. 기린의 목이 길어진 것은 목을 사용한 결과가 아니고 목이 긴 놈만이 살아남았기 때문이다. 이것이 다윈의 "자연선택"이다.
이 논리를 확장, 적용하기는 어렵지 않다. 표범이나 치타는 왜 모피에 점박이 얼룩 무늬를 갖고 있을까? 나뭇잎 사이로 빛이 산란될 때 대상은 어른 거리게 되는데 마치 얼룩무늬처럼 보인다. 그래서 얼룩무늬를 한 표범이 숲속으로 들어가면 주변 환경과 구분되지 않기 때문에 자신을 위장한 채 먹이감에 접근할 수 있다. 그런 놈만이 먹이감을 충분히 구할 수 있고 따라서 자신을 닮은 유전자를 후세에 퍼뜨릴 수 있을 것이다. 그래서 전투복도 적에 대해 자신을 은폐하기위해 얼룩무늬 문양이다. 빙고!

그림 1 고양이과 동물의 줄무늬
그러나 이러한 식의 답은 그럴듯하기는 하나 무엇인가 미심쩍스럽다. 점박이 얼룩무늬가 그렇게 좋다면 원숭이에게서는 왜 점박이를 찾을 수 없는가? 원숭이는 먹이를 사냥해야 하는 육식동물이 아니기 때문에? 그러나 얼룩무늬가 은폐술이라면 그것은 공격용인 만큼 수비용일 수도 있다. 얼룩무늬는 매의 매서운 눈에서 원숭이가 자신을 은폐하는데 아주 유용하다. 왜 원숭이는 얼룩무늬를 진화시키지 않았을까? 그것은 영장목에게는 얼룩무늬가 애당초 그 목의 설계도(유전자 풀) 속에 없었기 때문이다. 아무리 좋은 것이라해도 없는 연장을 불러낼 수는 없다.
소박한 진화론자들은 어떠한 것도 자연선택이 만들어낼 수 있다고 생각한다. 그러나 자연선택은 무한한 선택지의 팔렛트를 갖고 있는 것이 아니다. 각각의 생명체는 자신의 기본설계안에서 가능한 선택지를 가질 뿐이다. 점박이 얼룩무늬가 아무리 좋은 것이라 하더라도 그것은 영장류목의 팔렛트 안에는 없다. 똑같은 논리로 직립자세(영장목의 기본 설계안)가 아무리 좋은 것이라 하더라도 표범과 같은 고양이과 동물의 팔렛트 안에는 없다.
이것은 무엇을 의미하는 것인가? 형태형성을 이해하기 위해서 그 원인을 필요조건과 충분조건으로 주의깊게 구분해서 생각해 보아야 한다. 이 두 조건을 받아들이면 적응주의자와 구조주의자 간의 오랜 논쟁을 통합시킬 수 있다. 구조주의는 형태형성의 필요조건에 대한 논의이며 적응주의는 그 충분조건에 대한 논의이다. 자연선택에 의한 표범의 얼룩무늬의 설명은 충분조건이며 이것으로 설명이 완료되었다고 볼 수 없다. 왜 그것이 원숭이에게는 없으며 표범에게만 있는지를 설명하기위해서는 그 필요조건에 대한 탐색이 필요하다.
『판단력비판』에서의 칸트와 괴테, 셀링, 헥켈(Heckel)로 연결되는 독일의 자연철학, 『성장과 형태에 관해서』의 톰슨(D'Arcy Thompson)으로 연결되는 이 구조주의의 위대한 전통은 1950년대 앨런 튜링에 의해서 새로운 전기를 만났다. 구조주의가 기술(記述)을 넘어 설명(說明)의 단계로 넘어갈 수 있는 수단이 튜링에 의해서 제시된 것이다. 아래 글은 이 주제에 관한 Phillip Ball의 The Self-Made Tapestry 4장을 초역한 것이다.
1. 튜링의 반응-확산파
튜링은 튜링머신이라는 가상기계를 통해 컴퓨터의 기본논리를 제공한 수학자로서 우리에게 잘 알려져 있다. 그러나 그의 잘 알려져 있지 않지만 이 보다 결코 중요하지 않다고 할 수 없는 또 하나의 분야를 개척한 선구자이다. 그는 형태형성의 비밀을 밝혀 보고자 했다.
1952년에 그는 화합물의 최초의 동질성(uniformity)에서 자발적 대칭성 파괴가 일어나고 안정적 공간패턴이 발생하는 과정에 대한 가설적 화학반응을 기술하는 논문을 발표했다. 그는 여기서 최초의 수정난 의 대칭적 구형에서 어떻게 패턴화가 일어날 수 있는가에 대한 모델을 제시했다. "형태발생의 화학적 기초"(the chemical basis of morphogenesis)라는 이 논문은 이론생물학의 전 영역에서 가장 영향력있는 논문들중의 하나이다.
매질속으로 화학종(chemical species; 오늘날 모포겐morphogen이라 불린다)의 확산이 대칭성 파괴의 구동력이라는 튜링의 제안은 아주 중요한 것 중의 하나이다. 이것은 우리의 직관과는 어긋난다. 통상적으로는 확산은 동질성을 만들어내는 메카니즘이다. 즉 잉크방울은 물속에 고루 확산되면서 전체가 동질화된다. 그런데 튜링은 오히려 이 확산이 응집을 일으켜 잉크방울이 된다는 것이다. 그러나 튜링은 특별한 조건에서 이것이 일어날 수 있음을 밝히고 있다.
튜링의 화학계내에서는 확산은 또 다른 화학반응인 자기촉매적 과정과 경합해서 작용한다. 요컨대 이것은 반응-확산 시스템(reaction-diffusion system)의 사례이다. 튜링은 어떤 조건하에서는 여기된 반응-확산 시스템에서 안정적 패턴이 만들어질 수 있다는 것을 보여주었다. 이때 화학농도의 차이에 따라 띠의 형태가 만들어진다.
튜링은 어떤 화합물 예컨대 a가 자기자신을 더 많이 생산하기 위해서 자기촉매적 반응을 겪는 과정을 고찰했다. a가 생산되는 비율은 이미 존재하는 a의 양에 의존한다. 그러나 이 틀내에서 a는 또한 더 많은 a의 생성을 방해하는 화합물b의 형성을 활성화시킨다. 공간적 패턴을 형성하는 주요한 요인은 a와 b가 각기 다른 비율로 반응매질 전체로 확산되어 그들 각기의 영향의 범위가 상이하다는데 있다. 이것은 a와 b의 지배영역이 다르다는 것을 의미한다..
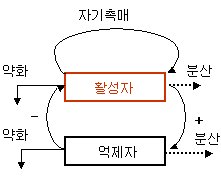
그림 2 활성-억제 모드 모식도
그러나 튜링이 화합물a의 활성적 기능(activation)과 화합물 b의 억제적 기능(inhibition) 간의 경합을 통해서 나오는 패턴발생의 메카니즘에 대한 중요한 해결책을 알게된 것은 자신의 논문이 출판된 이후였다. 패턴이 발생하기 위해서는 억제자 b가 더 신속하게 확산되어야만 한다. 그래서 a의 활성과 자기촉매적 생산은 국지적 과정이고, 반면 b에 의한 a의 제어는 넓은 범위에서 작용해야 한다. 왜냐하면 a근방에 b가 일단 형성된다면 이것은 다른 곳에서 a의 생성을 저지하기 위해서 신속히 확산될 수 있기 때문이다. 동시에 b의 신속한 확산은 a의 국지적 생성을 막지 않는다는 것을 보증한다. 전체틀은 활성-억제 시스템이라 불리는 반응확산 과정의 한 경우임을 보여준다.(그림2)
일단 a의 초기농도에서 무작위적인 요동이 자기촉매적 과정을 통해서 a의 증가로 반점이 만들어지기 시작하면 억제자 b가 생산되어 급격히 확산되면서 직접 인근에 활성자가 생성되는 것을 억제한다. 그래서 고립된 a의 반점의 열들이 b에 둘러싸이게 된다.(그림3 좌) 만일 반응의 생성물을 일정한 비율로 제거하고 a와 b를 만드는데 필요한 새로운 재료를 공급해 준다면 이 점의 패턴들은 장시간 동안 안정적으로 지속될 수 있다. 또 다른 형태는 a의 생산물의 영역이 띠의 형태로 되면서 미로같은 흔적을 만들어 낸다.(그림3 우)

그림 3 활성-억제 시스템은 점과 띠의 패턴을 만들어낸다.
그 뒤 1972년 마인하트와 기어르(H.Meihardt & A.Gierer)는 짧은 범위의 활성자와 긴 범위의 억제자가 튜링패턴의 중요한 원리라는 중요한 통찰을 얻어내었다. 그들은 더 복잡한 비선형방정식을 사용해서 튜링의 메카니즘을 정식화할 수 있었고 컴퓨터를 사용해서 그것이 실제 생물들의 패턴화하는 방법과 연관되어 있다는 것을 보여줄 수 있었다.
튜링패턴의 이 안정적 특성 때문에 이것을 그 반응의 최종산물로 간주하기 쉽다. 그러나 그렇지 않다. 그 패턴은 비평형적 패턴이며 동력학적 패턴이다.화합물의 혼합속에 끊임없는 운동과 반응에 의해서 유지되어 진다. 튜링패턴은 그 시스템이 평형에서 멀어지는 한 즉 대칭성이 파괴되는한 유지된다. 이것은 평형으로부터 멀어지는 구동력이 증가됨에 따라 대칭성이 파괴되는 불안정성안에서 동질적 매질로부터 자발적으로 생겨난다.
2. 띠무늬를 만드는 것
오늘날 이것은 화학적 패턴형성이라는 하나의 분야로 자리잡았는데 이것이 자리잡기에는 튜링의 논문 발표 후 20년을 기다려야 했다. 당시 형태형성을 연구하는 생물학자들 아무도 여기에 주목하지 않았다. 그러나 오늘날 튜링의 착상은 획기적인 것으로 받아들여지고 있다. 왜 그렇게 지체되었을까? 당시의 화학의 중요관심사는 이 비평형 현상 보다 평형 현상에 집중되어 있었다.
1960-70년대 화학자들이 벨로우소프-자보틴스키 반응(Belousov-Zabotinsky reaction)에 대해서 알게되면서 이 반응과 튜링의 확산 반응 도식간의 연관성이 지각되기 시작했다. 1971년 자보틴스키와 윈프리(A.Winfree)가 각각 독자적으로 BZ반응내에서 나선형 파를 관찰했을 때 더 구체화되었다. 이어서 윈프리는 이 나선파가 완전히 섞이지 않은 BZ혼합물에서 일어나는 활성자-억제자의 패턴형성 과정의 결과라는 것을 보여주었다. 이것은 화학반응에 있어서 비선형성, 자기촉매 그리고 되먹임 등에서 튜링의 구조와 유사하지만 그러나 BZ 화학적 파동을 발생시키는 메카니즘은 튜링구조를 만들어내는 불안정성과 같은 것이 아니다. BZ패턴은 확산파(travelling wave)인데 대해서 튜링의 패턴은 고정파(stationary wave)이다. 요컨대 국지적 활성과 넓은 범위에 걸친 억제는 그자체로는 안정된 튜링패턴을 만들어내기에는 충분하지 않다. 그것은 활성자의 농도의 변화에 대한 억제자의 반응이 신속할 때 (결과적으로 억제자를 배제하는 과정이 활성자를 배제하는 과정보다 더 빨라야 한다)만 생산될 것이다. 반면 억제자가 오랫동안 머무르고 있다면 그 시스템은 BZ와 같은 진동을 보일 것이다. 여러 화학종들간에 확산률에 차이가 없을 때 조차도 그러한 진동은 일어날 수 있다. 그러나 이 차이는 튜링 메카니즘에 있어서 본질적이다. 반면 확산파는 억제자가 활성자 보다 더 느리게 확산된다면 생성되어 진다. 그리고 이러한 파는 자발적인 전역적(global) 대칭성 파괴의 불안정성에서 발생하기 보다 매질에의 국지적 교란에 의해서 시발되어지는 것이 필요하다. 형태발생의 관점에서 볼 때 이 차이는 중요한 것인데 왜냐하면 튜링은 배에서 어떻게 안정적이고 고정된 구조가 자발적으로 생겨나는가를 설명하고자 하기 때문이다.
1968년 프리고진(Ilya Prigogine)은 진동반응에 대한 가설적 도식을 제안했다. 후에 브뤼셀레이터(Brusselator)라 불리게 된 이 모델은 BZ반응과 같은 진동과 분지(bifurcation)를 보여주었다.(그림4) 각종 화학종이 아주 다른 비율로 매질속으로 확산되어지는, 불완전하게 혼합된 계 내에서 진행되었을 때 그 불안정성이 튜링형의 안정적 반점과 띠를 형성하는 공간패턴- 부분적으로 다른 구성의 패턴 -을 발생시킨다.

그림 4 브뤼셀레이터 반응도식
이 가설적 브뤼셀레이터 도식은 튜링의 생각을 더 구체화시켰으며 BZ반응과 유사한 진동반응이 정확한 조건하에서 튜링패턴을 만들어낼 수 있다는 것을 보여주었다. 1990년대에 와서 이것이 실험적으로 입증되었다. CIMA라 불리는 이 반응은 드 케퍼(De Kepper)와 그 동료들에 의해서 1980년대 초 BZ반응에 대한 대안으로서 개발되었다. 그것의 진동과 패턴을 형성하는 거동을 보여주기위해 색깔을 변화시키는 지시제(indicator)를 부가했다. 그것이 반응에 관계하고 있는 요오드 3가 이온을 붙잡으면 노란색에서 푸른색으로 변한다. 이것은 여러 가지로 BZ반응과 유사하지만 튜링의 도식에 더 가까운데 왜냐하면 그것은 활성자와 억제자인 요오드이온과 염소이온을 각기 갖고 있기 때문이다.
활성자-억제자 시스템을 튜링구조를 형성할 수 있는 것으로 전환시키기 위해서 두 화학종이 아주 다른 속도로 반응매질 전체로 확산되어져야만 한다. 그러나 이러한 조율이 쉽지 않았고 그래서 오랫동안 그것을 실험할 수 없었다. 보르더 대학의 연구팀Bordeaux researchers들은 CIMA 반응을 고분자의 겔 속에서 대전帶電시킴으로써 화학종의 확산율을 임의로 조정할 수 있는 방법을 찾아내었다. 이것으로 해서 이제 이 실험이 가능해졌다.
다음 문제는 튜링패턴을 더 큰 영역위로 성장시키는 것이다.이것은 1991년 퀴양과 해리 스위니(Qi yang & H. Swinney)에 의해서 성취되었다. 튜링구조의 2차원적인 격자는 수천개의 노란색 반점들을 포함하고 있다. 겔의 온도가 18도 이상으로 올라가면 그 패턴은 사라졌다가 온도가 내려가면 새로 나타난다. 그들은 튜링의 예측의 또 다른 형태-반응조건을 변화시킴에 의해서 새로운 안정된 패턴을 형성할 수 있는 가능성-를 입증할 수 있었다. 요오드의 농도를 증가시키고 말론산의 농도를 낮춤으로써 새로운 방식으로 대칭성을 파괴하고 점대신에 띠를 형성할 수 있었다. 말하자면 조건의 변화가 표범을 호랑이로 바꾼 것이다.
3. 표피무늬의 문양
화학적 패턴형성에 관한 튜링의 아이디어가 통찰력있는 것이었음은 이제 의심의 여지가 없다. 그러나 이 모델이 형태형성과 적어도 어떤 관련을 갖고 있는가? 현재로서는 아직 확정적이지는 않다. 튜링의 불안정성과 흡사한 자발적 패턴형성 과정이 배발생의 가장 기본적인 문제라고 보고 있는 생물학자들도 있지만 대부분은 아직 테스트되지 못한 가설로서 보고 있다. 그러나 발달생물학 가운데 튜링의 아이디어가 부인할 수 없는 영향을 준 분야가 표범의 점, 얼룩말의 줄무늬, 기린의 반점 등 생물들의 표피무늬의 형성이다. 가장 인상적인 점은 무늬의 형태의 다양성에도 불구하고 같은 메카니즘에 의해서 설명될 수 있다는 것이다. 내 생각으로는 이것만으로도 튜링이 제대로 길을 찾았다고 볼 상당한 이유가 있다고 생각한다. 무늬의 세세한 것 까지 모두 유전자가 지시할 필요가 없으며 적절한 발달단계에서 활성자와 억제자를 만드는 청사진만 있으면 된다.
포유동물의 모피 패턴은 단지 몇가지 색의 모자이크이다. 흰색, 검은색, 갈색 또는 노란색/오렌지색 등 이러한 모피의 색이 어떻게 발현되는가는 잘 알려져 있다. 털의 색깔은 그 밑에 있는 색소분비세포인 멜라닌 세포에 의해서 결정된다. 멜라닌이라 불리는 색소는 멜라닌세포로부터 털로 통과하는 빛을 흡수하는 단백질이다. 거기에는 두가지 형태가 있는데 털을 검은색, 또는 갈색으로 변화시키는 오이멜라닌(eumelanin), 노란색이나 오렌지색으로 변화시키는 패오멜라닌(phaeomelanin)이 있다. 멜라닌 세포가 멜라닌을 분비하는지의 여부는 표피 바로 아래에 있는 어떤 화학물질의 현존 또는 결여에 의해서 결정된다. 그 화학물질이 무엇인지는 아직 잘 알려져 있지 않다. 제임스 머레이(J. Murray)는 이 화학적 '방아쇠'의 분포가 배발생의 첫 몇주 동안 튜링형의 활성자와 억제자의 상호작용을 통해 특징적 패턴을 형성하게 된다는 견해를 제시했다. 그래서 초기단계에서 배는 화학적 형태발생의 사전패턴을 획득한다. 그후 이것은 멜라닌세포에 의해서 읽혀진다. 이것은 마치 보통때는 보이지 않다가 촛불로 가열하면 보이는 잉크와 같다.
튜링적 불안정성을 일으키는 반응-확산 시스템에 의해서 생성된 시공패턴은 활성자와 억제자의 상대적 확산비율과 같은 요인들의 수에 의존한다. 패턴선택에 있어서 또 다른 강력한 영향은 크기와 형태이다. 사실 반응억제 시스템이 공간패턴을 형성할 수 있는 최소 크기가 있다. 패턴의 무늬의 특정적 크기-예컨대 반점의 반경-는 확산자의 확산율에 의해서 결정된다. 만일 전체 시스템이 이것과 크기가 같다면 어떤 패턴도 드러나지 않는다. 농도는 전체에 걸쳐 동일하다. 시스템이 성장함에 따라 지지될 수 있는 모드의 수가 증가함에 따라 끊임없이 복잡해 지는 패턴이 형성될 수 있다.(음향의 정상파와의 유비는 아주 인상적이다. 그림5 참조)

그림 5 활성-억제 패턴에 의해서 만들어지는 음파의 패턴
머레이는 크기와 형태에서의 이런 의존이 동물들의 꼬리에서 보여지는 패턴에서의 차이를 설득력있게 설명할 수 있는지를 탐구했다. 꼬리는 연구하기에 좋은 특징을 갖고 있는데 점점 가늘어지는 원통형으로서 수학적으로 근사하게 모델화할 수 있기 때문이다. 꼬리무늬는 두가지 기본패턴으로 되어 있는데 꼬리를 둘러싸는 줄의 형태이든지 아니면 점박이 무늬이다. 그러나 어떤 패턴이든 일련의 줄무늬로 끝난다.(그림6) 가늘어지는 원통형의 경우에 반응-확산 시스템이 어떤 패턴을 생성하는지를 계산했을 때 그 는 줄무늬와 점박이가 둘다 생성된다는 것을 발견했다. 만일 모델의 꼬리가 작다면 단지 줄무늬 만이 만들어진다. 이것은 단지 1차원적인 패턴인데 왜냐하면 색깔에서의 변형이 꼬리의 축을 따라서만 발생하기 때문이다. 만일 꼬리가 크다면 더 복잡한 형태들이 만들어지는데 축을 따라서 뿐 아니라 꼬리 둘레를 따라서 달라지는 2차원 패턴이 된다.(그림7) 그래서 꼬리 끝에서부터 넓어져 감에 따라 줄로부터 점박이 무늬에로의 변화가 치타나 표범에서 보듯이 일어난다.

그림 6 재규어의 아름다운 모피. 꼬리 말단으로 갈수록 점박이가 줄로 변한다.
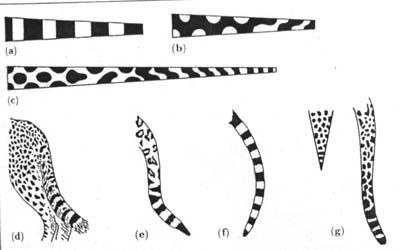
그림 7 꼬리의 둘레가 넓으면 점박이, 좁으면 줄무늬가 된다.a형은 사이즈가
작은 고양이에게서 b는 사이즈가 큰 표범이나 호랑이에게서 볼 수 있다.
머레이는 꼬리무늬의 종간의 차이도 그 종의 태아의 형태에서 설명될 수 있다는 것을 알아내었다. 사향 고양이의 꼬리는 전체가 줄무늬인데 대해 표범의 경우 꼬리 끝부분은 줄무늬이지만 대부분 주로 점박이 무늬이다. 성체의 꼬리의 형태로 보아서는 그렇게 될 이유가 없어 보이지만 태아의 형을 보면 그 이유를 알 수 있다. 태아 사향고양이의 꼬리는 가늘고 반경이 일정하다. 반면 표범 태아의 꼬리는 아주 짧고 끝이 가늘어지는 형태이다.
만일 성체 동물의 표피무늬가 어린 태아시의 화학적 사전 패턴에 따른다면 사전 패턴화단계의 시점은 아주 중요한데 왜냐하면 태아의 크기와 형태는 빨리 변하기 때문이다.이것은 얼룩말의 두 종류 Equus burchelli(a)와 Equus grevyi(b)의 띠무늬의 차이를 설명해준다. 전자의 띠무늬는 후자에 비해 폭이 더 넓고 그래서 그 수도 적다.(그림8 좌) 부르첼리가 그레비 보다 몇주 더 일찍 그 패턴을 형성한다는 것을 시사하는 증거가 있다. 동일한 화학적 메카니즘이 같은 넓이의 띠무늬를 만들면서 버첼리 태아에게 더 작은 수의 띠무늬를 줄 것이다.
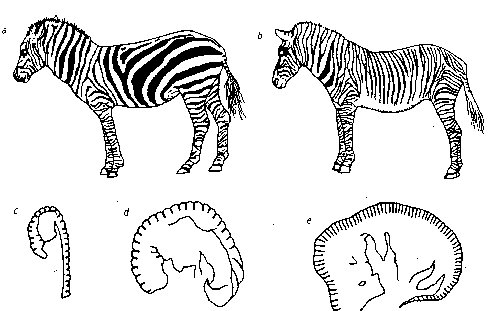
그림 8
여기서 한가지 주의를 덧붙인다. 띠무늬가 튜링유형의 모델에서 쉽게 만들어지는 전부가 아니다. 왜냐하면 띠무늬는 점박이로 부숴지는 경향이 있기 때문이다. 머레이는 그의 모델에서 띠무늬가 보존될 수 있기 위해서는 부가적인 요소가 일반적으로 필요하다고 가정했다. 예컨대 띠무늬는 활성자의 자기촉매적 생산율에 상한선이 있을 때 안정화될 수 있을 것이라는 것이다.
그림9

이 표피무늬는 scapular stripes라고 불리어 진다. 동체에서 띠무늬를 생성하고 다리에서 줄무늬를 생성하는 시스템이 동체와 다리의 교차점에서 갈매기 무늬를 생성한다는 것을 발견했다.
머레이의 모델내에서 반응-확산 시스템에서의 화학적 변수가 거의 모두 같다면 사전 패턴화의 시점에 태아의 크기와 형태는 그 후의 모피 패턴에 지배적 영향을 준다. 이것이 함축하는바는 회임기간이 짧은 작은 동물일수록 긴 큰 동물보다도 모피의 패턴이 단순할 것이라는 것이다.
그러나 패턴의 복잡성은 아주 큰 동물의 경우에도 오히려 줄어든다. 이것은 더욱더 많은 형태들이 가능해짐에 따라 무늬사이를 가르고 있는 선이 밀려나가면서 합쳐지기 때문이다. 예컨대 기린의 표피무늬에서 그 구분선은 아주 가늘고 코끼리나 하마의 경우 표지가 없다. 표피무늬는 복잡하면 오히려 단순해지는 말하자면 "과불급"이라 하겠다. 머레이 모델에서 기린의 패턴은 모서리가 둥근 물방울형태이다.(그림10)
그림10
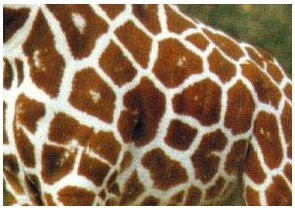
활성자의 생산은 어떤 임의의 장소에서 시작되는 것으로 가정했다. 활성자의 화학적 파동은 이 장소에서 퍼져나가면서 멜라닌 생산을 개시시킨다. 그래서 다른 파동을 만났을 때 서로 BZ반응처럼 서로 소멸한다. 이 소멸장소에서 활성자 생산의 각 영역간에 경계선이 만들어진다. 그래서 각 영역은 색소가 없는 영역으로 분리되어 멜라닌을 분비하는 다각형 영역들로 쪼개지게 된다. 이것은 실제 기린의 모피무늬에 대한 더 좋은 근사이다.
4. 패각과 같은 딱딱한 껍질에서..
동물의 모피의 무늬가 다윈의 선택압력에 의해서 결정된다고 믿고 있는 사람들은 상점에서 연체동물들의 패각들을 보면 놀랄 것이다. 이 패턴들은 아주 다양하고 아름다우면서도 어떤 분명한 목적의 쓰임새가 있는 것처럼 보이지 않는다. 그 현란한 무늬가 생존에 어떤 이점을 준단 말인가?
이런 패턴들은 확실히 유전적 조절을 통해 생겨난다. 그러나 그것이 선택압과 거의 관련이 없어 보이는 인상적인 생물학적 패턴들이 많다. 도대체 이 패턴이 무슨 기능을 하는지 다윈주의자들은 곤혹스럽겠지만 차라리 한스 마인하르트의 말처럼 자연의 "놀이"가 아닐까?
셀shell의 패턴을 포유동물의 점박이무늬나 띠무늬와 유사한 것으로 보고싶은 유혹이 생길 것이다. 사실 전체적인 2차원적 표피의 패턴화 과정에서는 분명히 유사한 점이 있다. 그러나 셀이 성장함에 따라 연속적으로 일어나는 과정의 역사적 기록을 나타내고 있다는 점에서 포유류의 그것과는 다르다. 왜냐하면 셀은 석회화된 물질을 가장자리에 계속 부착시켜감으로 성장하기 때문에 우리가 셀의 표면을 통해서 보는 패턴은 셀의 가장자리의 1차원 선을 따라서 색소가 분비되어서 만들어진 것이다. 따라서 성장축을 따라서 또는 그 둘레를 돌고 있는 띠무늬는 그것과는 질적으로 다른 동결된 시간의 역사이다. 가장자리의 주기적인 패턴은 셀이 성장해도 그 장소에 남아있다. 색소는 성장선 전체에서 일양하게 분비되는데 그것에 색소가 없는 성장주기가 따라 나온다. 성장방향에 대해서 비스듬한 각도로 달리고 있는 띠무늬는 셀이 성장함에 따라 가장자리를 따라 진행하는 색소 확산파의 표현이다.(그림11 우)
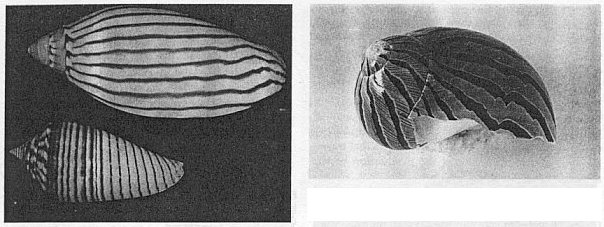
그림 11 (좌) 셀의 축에 수직인 띠와 수평인 띠는 아주 다른 패턴의 메카니즘을 반영하고 있다. 상단은 공간적으로 일양하지만 시간적으로 주기적인 패턴과정을 반영하고 있으며 하단은 그 역이다. (우) 비스듬한 사선의 띠는 시간과 공간에 대해서 주기적인 성장선growth edge에서의 이행파travellimg wave의 결과이다.
셀의 패턴은 튜링패턴과 유사한 정지패턴과 BZ반응과 유사한 확산파를 일차원상에 표현한 것이다. 그림 11 좌하단의 셀의 패턴은 1차원의 간단한 주기적인 정지패턴이다.(그림12 좌) 줄무늬의 넓이와 무늬간의 간격은 모델의 변수, 특히 활성자와 억제자의 상대적 확산율에 따라 민감하게 조율된다.(그림12 중간과 우) 같은 종간의 차이는 상이한 성장조건의 결과일지 모른다. 한편 종간의 차이는 셀의 가장자리의 성장부가 발생의 초기단계에 동결된다면 그래서 확산하는 화학물질을 통한 교신이 중단된다면 발생할 수 있다. 셀의 패턴은 가장자리 성장부의 패턴의 시간적 흔적이기 때문에 완전한 2차원패턴은 가장자리가 어떻게 성장했는가에 의존한다. 예컨대 그림11의 줄무늬 패턴은 성장선에서의 동일한 종류의 공간패턴의 결과이다.(그림11 좌 참조)

그림 12 셀의 성장선에 수직인 띠는 성장선에서 1차원 공간 패턴화의 결과이다. 셀의 성장선이 확장됨에 따라 띠의 패턴이 만들어진다.(좌) 확산이 너무 빠르게 일어난다면 띠의 폭이 넓어지고(중간) 활성자의 농도가 포화상태에 까지 오르면 띠 간의 간격이 불규칙해진다.(우)

그림 13 셀의 패턴의 1차원 세포자동자에 의한 시뮬레이션
5. 그러나 실제로 그런가?
그림14
학명 Pomacanthus
semicirculatus의
엔젤피시 
현장의 생물학자들은 미심쩍어 한다. 이 반응-확산 모델에 의해서 만들어진 패턴과 실제의 것 간의 유사성이 아무리 인상적이라 하더라도 그것은 단순한 우연의 일치일지도 모른다. 튜링의 메카니즘이 실제로 생명체에 작동하고 있다고 어떻게 확신할 수 있는가?
이 증명은 형태발현물질 morphogen의 발견에 있지만 이것은 아직 찾지 못하고 있다. 그러나 1995년 일본의 생물학자 시게루 곤도 Kondo 등은 앤젤피시angelfish의 표피무늬에서 튜링 메카니즘을 지지하는 증거를 찾아내었다. 이것은 반응-확산 시스템의 일반적인 결과이지만 지금까지와는 다른 점은 이 띠가 발생 초기 단계에 고정되는 것이 아니라 성장해 감에 따라 변해간다는 것이다. 성장하더라도 패턴에는 변화가 없으며 더 작은 물고기는 더 적은 수의 띠를 가진다. 예컨대 Pomacanthus semicirculatus라는 학명의 엔젤피시의 새끼의 줄무늬는 2cm이하이고 3개의 띠를 갖고 있다. 성장해가면 띠는 점점 두꺼워지고 4cm에 달하면 새로운 띠가 본래의 띠들 사이에 생겨나는 돌발적 변화가 일어난다. 새로운 띠는 본래의 띠들 사이에 생겨나며 띠 사이의 간격은2cm 이하의 어린 것에서 보이는 간격으로 좁혀진다. 이 과정은 8-9cm로 성장할 때 또 되풀이된다. 이와반대로 기린은 커지면 부풀어 오른 풍선처럼 패턴 자체가 커진다.
이것은 엔젤피시의 띠는 성장 과정 동안 활성 상태를 유지해야 된다는 것을 의미한다. 즉 반응-확산 과정이 계속 작동하고 있어야 하는 것이다. 만일 이 고기가 축구공의 크기 정도로 커질 수 있다면 짐 머레이의 작업에서 드러났던 사이즈이 효과가 작동해서 아주 다른 패턴으로 변했을지도 모른다. 그러나 물고기는 이 점에 도달하기 훨씬 전에 성장하기를 그친다.
곤도는 성장하는 셀의 열에서 발생하는 반응-확산 과정의 이론적 모델로 이 거동을 재현할 수 있었다. 이것은 단지 동일한 과정이 단순히 동물의 모피에서 보여지는 정지패턴을 만들어내는 것과는 달리 성장을 유도하는 패턴의 팽창을 만들어낼 수 있다는 것을 보여준 것이 아니다. 이것은 튜링 메카니즘에 대한 아주 설득력있는 증거이다.
그러나 연구자들은 한걸음 더 나아갔다. 그들은 아주 다른 줄무늬를 가진 Pomacanthus imperator을 연구했다. 이것은 어릴 때는 Pomacanthus semicirculatus와 마찬가지로 성장함에 따라 띠의 수가 증가하는 방식으로 자란다. 그러나 성체가 되면 띠는 물고기의 머리-꼬리 축에 평행하도록 조율된다. 그래서 그 띠의 수는 몸의 크기에 비례하고 띠 사이의 간격은 일정하게 된다. 띠에서 새로운 가지가 생겨나서 자라다가 분리된다. (그림15 좌상) 이 Pomacanthus semicirculatus에 대해 동일한 반응-확산 모델을 사용해서 곤도는 이 거동을 정확히 재현할 수 있었다.(그림15 좌하) 또한 그의 이 모델은 등과 복부 지점(고기의 위와 아래)에 위치한 분지점의 더 복잡한 거동을 시늉낼 수 있었다.(그림15 우)
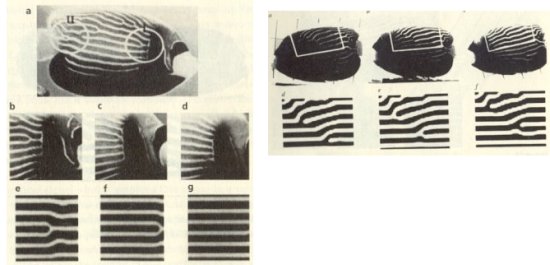
그림 15 엔젤피시의 띠무늬의 성장. 하단은 튜링 모델을 이용한 시뮬레이션
이 반응-확산 모델이 복잡한 물고기의 띠무늬를 재현하는 것을 보면서 이것이 자연과는 상관없는 것이라고 생각하기는 어렵다. 곤도는 이 반응-확산 과정은 표면적으로는 성체의 물고기에만 적용되는 것처럼 보이지만 이것은 단지 이 경우에 활성분자와 억제분자의 화학종을 확인하기가 쉽기 때문이라는 점을 지적했다. 이것은 자연이 패턴을 어떻게 만들어내는가에 대해 튜링의 추측이 타당하다는 움직일 수 없는 증명을 제공해 주고 있다.
6. 나비의 날개
지금까지 살펴본 것은 단지 두가지 색으로 된 동물의 표피무늬였다. 여기서 세포들에 의해서 생산되는 색소는 1가지 뿐이다. 그러나 자연이 연출하는 현란한 색깔들은 어슬픈 이론적 설명들을 압도한다. 한 예로 색의 만화경을 보여주는 나비의 날개를 보라. 색의 종류가 환상적으로 많을 뿐 아니라 그 무늬도 아주 정교하다. 사실 정교성에서는 기린의 표피무늬는 이것을 따라오지 못한다. 두 날개의 무늬가 아주 대칭적이어서 점과 띠들은 마치 정확히 측정한 다음 붓으로 그려 놓은 것 같다. 이 디자인이 어떻게 그려진 것일까?
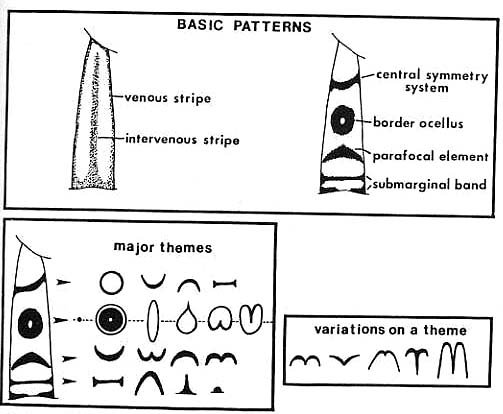
일찍이 1920년대에 슈반위취(Schwanwitch)와 쥐페르트(Suffert)가 이 문제를 가지고 씨름했다. 그들은 나비의 날개문양의 엄청난 다양성을 하나의 통일된 도식으로 종합했는데 이것이 "님팔리드 기본 설계안"(nymphalid ground plan)이라고 불리는 것이다. 이것은 나비의 날개무늬들에 들어있는 공통된 패턴으로서 자연에서 보여지는 엄청나게 많은 무늬들은 이 기본 청사진에서 어떤 것을 선택하고, 어떤 것을 배제하고, 또 어떤 것을 변형시킴으로써 만들어진 것이다. 이 둘은 각기 독립적으로 이 도식을 개발했지만 아주 놀랄 정도로 일치한다.(그림16)
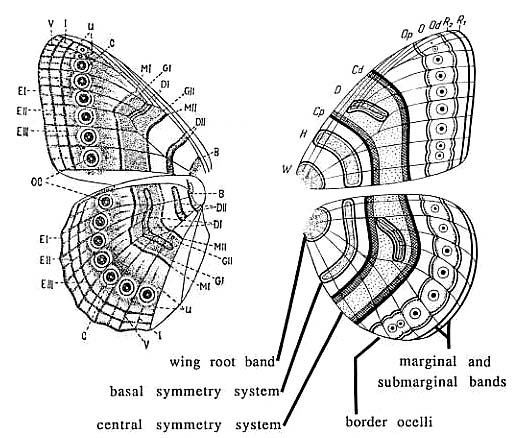
그림 16 nymphalid ground plan 좌측은 슈반위취(Schwanwitch)의 도식, 우측은 쥐페르트(Suffert)의 도식

기본 문양은 위에서 아래로 앞에서 뒤로 날개를 횡단하고 있는 점과 원호 그리고 띠들이다. 상하축의 문양(top-to-bottom)은 대칭 시스템이라고 불리워지는데 중앙의 대칭축을따라서 근사적인 거울상( mirror image)을 이루고 있기 때문이다. 아주 복잡한 문양이라 하더라도 쪼개보면 두세개의 대칭 패턴들의 조합으로 환원할 수 있다.
이 문양을 만들어내는 기본 단위는 지붕의 기와처럼 날개를 덮고 있는 아주 작은 비늘(鱗片)이다.(그림17) 이 비늘들은 각기 하나의 색으로 되어 있는데 확대해보면 텔레비전의 픽셀처럼 보인다. 이 색들은 멜라닌 -동물의 표피무늬를 만드는 색소- 이라 불리는 화학 색소에 의해서 만들어진다. 흰색, 빨강색, 노란색 등이 그것이다. 그러나 진짜 미묘한 색깔은 화학이 만들어내는 것이 아니고 물리학이 만들어내는 것이다. 비늘의 융기된 주름(rib)에 빛이 반사하거나 산란하면서 현란한 색이 만들어지는데 초록색과 푸른색은 이렇게 해서 만들어진다. 물론 이 님팔리드 기본 설계안의 모든 요소들을 다 갖고 있는 실제의 나비는 없다. 이 기본안은 자연이 사용하는 가능한 모든 문양들의 총합이다. 크기, 형태, 그리고 선택된 요소들에 부여하는 색상 등을 이리저리 조립함으로써 어떤 문양이라도 이 기본안에서 만들어낼 수 있다.(그림18)
그림 17 (좌) 나비의 날개를 확대한 것. 미세한 비늘들이 모여 만들어진다.
그림 18 나비의 날개 디자인의 보편적 도구상자.